| |
|||
|
Related Products Astronomical Society of the Pacific
|
![]() A Biologist's View of Life Out There
A Biologist's View of Life Out There
sponsored by

|
A Biologist's View on Life Out There A multitude of stars in the Galaxy, and increasingly it looks as if many of them harbor worlds. But what of life on those worlds? First we must consider the natue of life on our one. by John M. Aguiar Reprinted from Mercury, Mar.-Apr. 1999. "Many biologists," George Musser, former Mercury editor, wrote me over a year ago, "have long been on the pessimistic side of the SETI discussion, and astronomers on the optimistic side." The differences between these two professional groups all boil down to immensity. For an astronomer, a galaxy with some two hundred billion stars will offer myriad opportunities for even the wildest chances to crop up. A biologist, meanwhile, surveys the tens of millions of species on Earth–only one of which has developed electromagnetic technology–and concludes that intelligent life must be supremely scarce. I am caught in between: a biologist by love and training, an amateur astronomer by irresistible draw. I read Cosmos and Jacques Cousteau side-by-side as a boy, and it never occurred to me that they should one day be pulled into different worlds. Exobiology is the natural crux of these two disciplines, their point of ultimate interdependence; yet its central assumption, that life is not isolated to our one world, is championed by astronomers more vigorously than biologists themselves. In casual conversations, those in the life sciences will admit the possibility; on listservs one finds a receptivity, even an enthusiasm for the idea of other life–if not perhaps other intelligence. But in formal publications, some prominent biologists have starkly disavowed all likelihood of contact with other species, pointing up the vast improbability of our own genesis and castigating SETI as a waste of resources. Distant
Interest But despite the recurring announcements of newly discovered planets–proof of a critical prerequisite–most in the life sciences still consider the topic irrelevant to their professional lives, fit only for engaging speculation. Unlike astronomers, constrained by distance and serendipity, biologists are accustomed to working in direct contact with their subjects (happily knee-deep, sometimes) in an atmosphere where no point is proven without manipulation. Even if spectral analysis of a nearby star suggested a living world, not even the planned NGST (see "Astronomer's Notebook," Jan/Feb, p. 8) could resolve its surface features; and the vastness of interstellar space would mean that even a remote surveyor moving at unprecedented speed, such as Robert Forward's Starwisp, would not be likely to arrive within the lifetime of today's researchers. The return of samples would necessarily be an issue of centuries. With such marginal chances for personal involvement, and their pervasive disinterest in extrasolar life at large, biologists tend to be unimpressed with SETI's potential value. But there are those few–notably the Nobel laureate Christian de Duve–who are more sanguine on the subject. "Life and mind appear as cosmic imperatives," asserts de Duve, "written into the fabric of the Universe. Given the opportunity, matter must give rise to life, and life to mind." Unlike those who criticize SETI from a background in paleontology, de Duve's many decades of experience with cellular processes give him an intimate feel for the chemical basis of life. Most biologists regard life's appearance as an event of stunning improbability: a repetition, they feel, is simply too unlikely. Their arguments are grounded in assumptions of contingency, the ultimate sway that random events are believed to hold over the shaping of organisms (and ecosystems) in evolutionary time. From this perspective, the origin of life on Earth is asserted to have been the result of pure chance–just enough happy accidents of chemistry, with no expectation it could ever happen again. De Duve refutes this; for life to have arisen here through purely chemical agencies, he argues, the steps leading to it "must have been strongly deterministic and reproducible," and should recur throughout the Universe wherever favorable conditions obtain. Where the conditions are right, life not only can occur, it must occur.
Some
Assembly Required But would favorable conditions be likely to obtain? The most critical, for our sort of life, is the presence of water: not only liquid water, required as matrix by our cells, but also the ice and water vapor necessary to the complex global cycles of a living planet. Thus the presence of water at tripoint is doubly vital, both for lifeforms and the world that sustains them. This would depend on the nature of the primordial gas and dust that form the swaddling clothes of a world, as well as on impacts from other nascent bodies that might drive off intrinsic moisture. Yet if we continue with the neutral assumption that our Sun is aggressively average among its spectral peers, and allow that such Moon-forming collisions as ours are not common, planets bearing sufficient liquid water become more than plausible. Improbably near to us, evidence piles on evidence for a global ocean beneath Europa's mantling ice, warmed by gravitational flexing from Jupiter and its other giant moons. If two waterworlds can be found in a single system, it says a great deal for the prevalence of liquid water elsewhere in the Galaxy. The orbital spread of the giant planets detected to date tends to fall well within the 1 AU range, but this only expands our understanding of what configurations are possible. Smaller bodies must whirl unseen in other orbits; the question becomes, "Which distances are appropriate for life?" The answer: "It depends." Water can only exist at tripoint in a narrow torus around a star, the "water trough" or ecozone. The diameter of the ecozone is unique to each parent star, determined by its mass and age on the main sequence. But if planets are likely to have formed around many of the candidate stars, it strongly suggests a population of ecozone worlds forming through random assortment alone, if not by some more ordered process. Even those systems with gas giants shouldering close to the star may harbor life, if those giants possess equally Brobdingnagian moons. The Galileo spacecraft's recent discovery of a magnetic field around Ganymede reinforces the potential, on other massive moons, for a biosphere similar to our own. The magnetosphere serves as a shield against the naked energies of its parent star, and would also protect an atmosphere from the erosive radiation of its patron giant. Even with water and a protective shield, other needs must be met. Would the requisite elements be available for electrochemical life such as our own? Hydrogen, carbon, nitrogen, and oxygen are the "Fab Four," essential to our basic design; but there are also those elements ("metals" to astronomers, "nutrients" to biologists) vital to specific molecular structures: potassium, phosphorus, calcium, sulfur, and magnesium, as well as trace elements including iron, zinc, boron, lithium, manganese, molybdenum, chlorine, and copper–not to mention the more exotic oddities, such as vanadium or cobalt, found in tunicates and other specialized organisms. The metallicity of a star (its richness in elements heavier than hydrogen and helium) would clearly exert a subtle but critical effect on the biochemistry of dependent worlds. While a star's metallicity is dependent on its formative milieu, those stars rich in potential nutrients would tend to pull them close during the early stages of solar-system formation: denser elements are generally better able to survive the heat near the nascent star, while lighter ones are blown outwards by the protostar's intensifying radiation. The result of this gradient would be a concentration of life-sustaining elements in the shallows of the star, overlaid on the ecozone in a warmwater matrix. This density gradient recurs in miniature around gas giants, seen most clearly in the Galilean Moons of Jupiter: sulfur-heavy Io skimming the cloudtops, icy-light Callisto furthest of the four.
Biology
of Numbers Assigning failure to a species is problematic on its own. One species may diversify into several that persist beyond the original form; that first species has no more "failed" than a parent whose children carry on–and in fact, a diversity of species is considered the essence of success for a lineage. It is the failure of lineages, with all their plethora of species, to produce frequent intelligence that is a more challenging problem. In the continuity of life, extinction is foiled by the radiation of new species; in the history of our biosphere, both diversity (the number of species) and biomass (their sheer substance) have burgeoned through time. But lineages–great clans of related groups, such as mollusks, or vertebrates, or crustaceans–are another matter: The trilobites, an ancient group of marine arthropods, are the best-known of several major groups which have entirely vanished. The apparently random termination of entire phyla underpins those arguments which support pure chance as the overriding force in all evolutionary history. It is by this same randomness, on the individual level, that mutations create variability–the spread of individual differences within a population, upon which natural selection acts either to cull or magnify a character, such as fur color or molar size. This is fundamental to evolutionary theory; but Christian de Duve points out that contingency has been made to shoulder more than it should carry. He cautions against the misconception that evolution skitters along at mutations' whim. Rather, mutations more often reside quietly in a population's collective gene pool, awaiting the moment when conditions change and they may become the key to survival. And there are other forces at work. "Chance does not exclude inevitability," de Duve writes. In his view, for instance, the advantages of interlinked neurons lead to the selection of greater neural complexity, no matter the body plan. This is borne out today in the brains that govern the radically different body plans of octopi, cetaceans, and anthropoid apes–and our world's first taste of this complexity appeared with eruptive force.
Life's
Own Nova Perhaps the clearest example of this is the evolution of flight, which has arisen no less than four separate times in the past three hundred million years–once in the insects and thrice more from a single subphylum, the vertebrates. The difference between insect wings and those of the later innovators (pterosaurs, birds, and bats) is dramatic enough: relatively stiff, flat sheets with only one point of articulation for insects, as opposed to the flexible, multi-jointed appendages of vertebrate airfoils. But even these three sets of kindred flyers are entirely distinct from one another, with bone and musculature unique to each group– not to mention feathers. Even more remarkably, three of the world's four great flying clans converged not only on level cruising, but also on a far more complex and difficult maneuver: the hover. Hummingbirds, nectar-feeding bats, and uncountable insects are all capable of hovering at will, each group adapting its own prior solutions to the same aerodynamic problem. More specific parallels abound. Today's flamingos are long-legged wading birds that snake their heads across shallow water, sieving out zooplankton (such as tiny shrimp) with fibrous strainers in their beaks. Some 140 million years ago, a pterodactyl now called Pterodaustro also sieved shallow water through specialized strainers, and probably for much the same sort of prey. (And since pigments from the shrimp they eat give modern flamingos their color, we must soberly consider the prospect of pink pterodactyls.) Nor is this tactic limited to flying forms: mysticete whales use immense strainers of keratin fiber, collectively known as baleen, to skim up larger zooplankton.
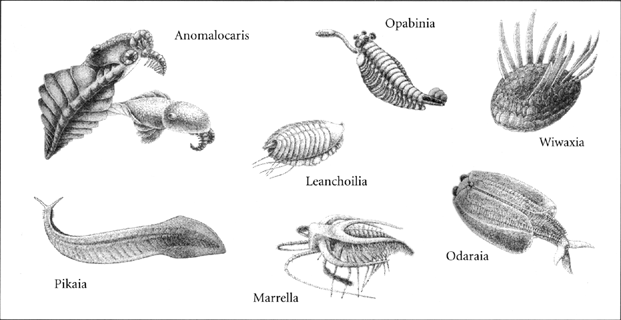
So while each species is unique, many have followed their own paths to arrive, if not at precisely the same form, at least at a configuration that gets the same job done. Nor need these pathways be obvious, even in hindsight. An unobtrusive shape among the bizarrely festooned creatures of the Cambrian was the undulating, pinky-length Pikaia, long considered the oldest identifiable chordate until supplanted by the slightly older Yunnanozoon. As a progenitor to the lineage that eventually gave rise to tool-users, Pikaia seems an unlikely candidate when compared to Anomalocaris or Opabinia, which at least had grasping appendages. But from Pikaia and Yunnanozoon (or creatures very much like them) unfolded a wellspring of life that produced hatchetfish and hummingbirds, lemurs and elasmosaurs, glass frogs and humpback whales. Rather than a warning of near-certain doom, the Cambrian jubilee demonstrates the array of forms a biosphere may forge from a single lineage, amazingly malleable beneath the hammerblows of natural selection. All
the World's a Stage Each species represents a solution, in a sense, to the problems posed to organisms by their physical and biological environment. But as in mathematics, there is often more than one pathway to a solution, and the process is endlessly iterative. Each new solution–each new species–will change its environment in some way. A mangrove seedling sends down roots on an empty tidal flat, creating a pioneer colony of trees in shallow water; their tangled roots will trap silt from the water and eventually build up new land. A new predator arrives on an island, devastates a previously unchecked herbivore; plants that would have been eaten as seedlings now rise to maturity, and the shade of their growth affects other species in turn. Each new change poses problems to other members of the community, who must meet the challenge or perish. And beyond all this lie the abiotic factors, the details of rainfall and soil type, glaciation and seafloor spreading, that would be specific to each world. All these factors exert pressures on native organisms, who both create and endure their overall environment. There are limits; there is fierce competition for resources–but also cooperation: commensalism and mutualism, species evolving in concert as symbionts of various stripes, the innovations adopted because they work, on other worlds as well as our own. Each biosphere would follow its own trajectory, creating patterns with unique detail–yet traced by a familiar outline. None of this, on its own, creates intelligence. Tools
of the Trade
Nor are we alone in this regard. Mastery of tool use was long considered an exclusively human trait; we are only gradually understanding its true extent among animals. One skeptic of SETI allowed that chimpanzees "and a few others" are also tool users, adding that they "are not very successful." Few hardly describes the growing number of species known to use tools–that is, to manipulate objects external to the body toward a preselected goal. (The concept of forethought, necessary to discussions of tool use, is itself a challenge to the dogma surrounding animal cognition.) Jane Goodall's trailblazing studies of chimpanzees first brought tool-use in animals to the fore, though the other three species of great ape–bonobos, gorillas, and orangutans–show less aptitude in the wild. We now recognize a wide range of tool-users from among other vertebrates, including baboons, macaques, capuchins, elephants, dolphins, sea otters, a variety of birds–jays, crows, marshtits, nuthatches, and finches–and even invertebrates such as octopods and several species of ants. There are no bandsaws or electric drills here, but the objects employed range from sticks, stones, and sponges to complexly fashioned hooks–a diversity that suggests greater mental potential than had been conveniently assumed. This collective toolkit reflects what must be a fundamental potential for tool use in ecosystems around the world. Humans are not an anomaly that hit on this trick alone; we have only elaborated a strategy common to a surprising breadth of animal life. Few of these species approach the chimpanzees (Pan troglodytes) for the complexity of their tool use and the thought behind it; the comment that they are "not very successful" is a slight to their achievements. Like humans, different chimpanzee populations pass on unique cultural traditions. While the chimps of the Gombe Valley in Tanzania have been studied the longest, beginning with Jane Goodall nearly forty years ago, it is the chimps of the Ta'i Forest in C6te d'Ivoire, across the breadth of Africa, who have the richest repertoire of tool-use. In addition to the use of tools for water sponging, honey-fishing, ant- and termite-dipping and a variety of threat displays (all of which are known from chimps in Gombe and nearby Mahale as well), the Tai chimps also use hammers, both wooden and stone, to crack open hard Panda nuts, carefully arranged on natural stone anvils. The Swiss ethologists Christophe and Hedwige Boesch have shown that the Ta'i chimps can fix the position of various tools and anvils in mental maps, direct equivalents to the Euclidean maps developed by human minds in childhood. The Ta'i chimps use these advanced spatial constructs to navigate through dense rainforest, and they keep track of particular tools for years at a time. (This is a better record than most suburban humans can boast.) To their immediate west in Guinea, other chimpanzees wield palm fronds in a mortar-and-pestle arrangement to extract palm oil, and in neighboring Sierra Leone still other chimps use a primitive sort of footwear when clambering through spiny kapok trees. It was in Sierra Leone, four hundred years ago, that Portuguese-African adventurers first reported these and other chimpanzee behaviors; their accounts were dismissed as fantastic embroidery until recently, when many of the anecdotes have been verified by field research. But pounding nuts is a long way from crafting radio telescopes, and all of the above might be glossed as irrelevant because none of these species, not even chimpanzees, are working with microchips. There is a narrow biological truth to the remark that chimps, despite their talents, are not especially successful: restricted to remnant pockets of rainforest and savanna, isolated and in danger of extinction. But instead of a curious footnote, their tool-using abilities should be recognized as intrinsic to the basal stock from which our own ancestors arose. Walk
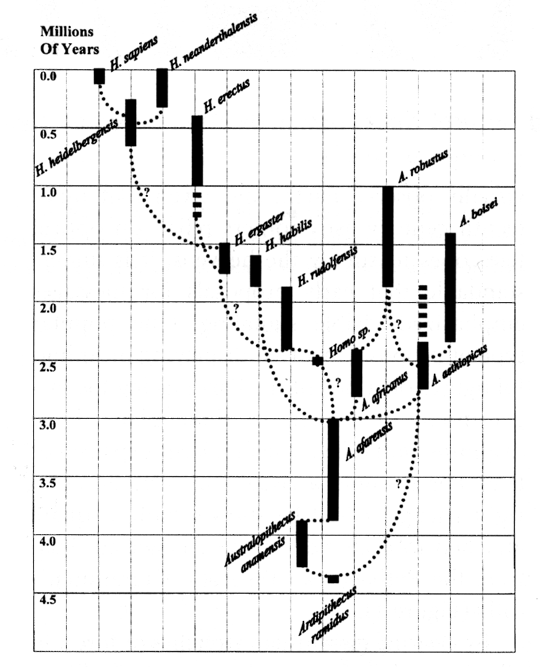
On
What, then, led to our one species developing technology beyond stone hammers? The defining feature of our small clan: bipedalism, adaptation to the wide-open in place of the densely forested. Humans are unique among primates not for tool use nor politics nor expression of culture, but for a phenomenal range, symbolic language, and the now-casual mastery of fire. The latter two are predicated on the first. A handful of primate species brave snowy mountains or desert fringe, but the great majority are confined to tropical forests, now shrinking worldwide beneath the fires and saws of our own species. Our global distribution gave us the resource base to support increasingly advanced technology, as well as a wide enough range of populations to develop it. But we were not the first hominids to use tools or to spread across the continents. Accumulating evidence, including a report in Science from March 1998, suggests that Homo erectus, a hominid closely related to ourselves, had a rudimentary grasp of seafaring nearly a million years ago. H. erectus–along with H. heidelbergensis, which may have given rise to ourselves and the Neandertals, H. neanderthalensis–represent the most recent bunching of hominids in a larger cluster extending back at least four million years, a clan deeply rooted in Africa and including as many as fifteen separate species. Among them are names that have been familiar for much of the century, such as Australopithecus africanus and Homo habilis, and new species like Ardipithecus ramidus, only described in 1995. While brain size varied widely among them, they all had an easy upright stride, and their distribution must have been wide, if not pancontinental. Bipedalism, the hallmark of the Hominidae, lends itself to wandering. Nor did each species explore a continent devoid of relations. Fossil findings suggest that between 1.5 and 2.5 million years ago at least four hominid species coexisted in Africa, perhaps more; how they interacted, and why one did not swiftly outcompete the others, remain unanswered questions. Hominids are usually considered to be consummate generalists, species that occupy so broad a niche (their physical and energetic position in an ecosystem) as to admit no competition. Similar species pursuing the same resources must specialize, subdividing their needs between them, or (so theory insists) one must become locally extinct. Enough local extinctions and the species as a whole will perish. A few hominid species, the "robusts," do indeed seem to have specialized: Australopithecus robustus, A. boisei, and A. aethiopicus all had the massive molars and chewing muscles that imply an exclusively vegetarian diet. Yet two or more species of Homo, with somewhat larger brains than the australopithecines, appear to have remained generalists–and somehow coexisted. We have no hard evidence as to the nature of that coexistence, or that between other hominid groupings; their encounters may have ranged, from instant hostility to long-term networks of trade. But the outcome of this ancient African milieu–ourselves, a single hominid species, lone survivors of the crowded group–poses a necessary thought: In the rough-and-tumble of species competition, did we survive because we were more violent? Or because we were more intelligent? Or worse: because of both? These questions are crucial for any consideration of the potential number of intelligent species in the Galaxy. Our hominid lineage, currently known as far back as Ardipithecus ramidus, seems to have met conditions that encouraged (or forced) diversification into several forms, including the vegetarian robusts. While we have no evidence of tool use from these or other australopithecines–as opposed to the abundant evidence for Homo habilis and later members of our genus–this is less a reflection on their abilities than a bias in the fossil record toward stone implements. Of all the tools devised by chimpanzees, only their stone hammers are durable, and these unshaped. The tools they do manufacture, by modifying existing objects, are all wood or bone: poor candidates for preservation, and not obviously tools unless witnessed. Given the abilities of chimps today, we may give the australopithecines the benefit of the doubt as potential tool-users; if competition forced their extinction, their lack of tools should not have been the issue. With this in mind we may speculate from one of two perspectives: Either the large cluster of hominid species was a necessary crucible, viewed retrospectively, for our own development–the competition between species driving physical, mental, and cultural advancements that saw the extinction of all species save our own forebears–or, alternatively, the other species in the cluster fell victim to changing environmental conditions which our ancestors alone could survive. Assuming this latter prospect opens two related possibilities: Our current position as sole surviving hominid may represent the exception rather than the rule (the vagaries of climate differing from world to world)–and, more hopefully, intelligent species may not be automatically lethal to one another. The implication is that rather than interspecies warfare being a prerequisite for the development of one technologically capable intelligence, a cluster of closely related species may evolve simultaneously, perhaps alternating between competition and cooperation in undestructive patterns. (The two most recent hominid species, ourselves and the Neandertals, apparently coexisted in the Middle East for at least ten thousand years–far longer than required for either to simply outcompete the other.) Different clusters of species, in turn, might arise at different periods in a world's history, as great lineages rise and fall through geological time. Rather than one world birthing a single intelligent species, we could expect its biosphere to produce clans or clusters of species, which–if they developed advanced technology alongside each other or cooperatively–might survive to disperse into the Galaxy, either severally or as a united multispecies culture. Once away from their birth-world, either continually starfaring or settling other planets, there is no reason why different selective pressures would not continue to generate new forms. During their time in the ecozone, some living worlds may yield up two or three successive clusters of species, shed like spores into each spring of a galactic year. Braving the Silence So where are they? Unlike many critics of SETI, I am not especially bothered by the lack of instant Galactic attention to our faint and jumbled radio traffic. It is only a vestige of earlier geocentrism that still expects the rest of the Galaxy to come running and marvel the moment we reach out, ever so fitfully, into space. That civilizations with long technological histories would communicate by our favorite methods is also unlikely. We may be leaking smoke signals, in effect, while the rest of the Galaxy is on email. Our presence may well have been noted by the equivalent of ham radio operators, hobbyists with an unofficial interest; they may respond in their own time. We should remain attentive for such a call, and develop new technologies to open our proxy ears to other possibilities. When passive listening is our only form of exploration, we must hew to it ever the stronger. But we cannot remain passive forever. The only thing left us to do–the only sure way to capture the attention of onlookers and reach for true maturity–is to slip anchor from the shallows of our star, and venture the greater ocean. JOHN M. AGUIAR is a graduate student in conservation biology at Texas A&M University. Between Star Wars and Carl Sagan he was hooked from an early age on the idea of a populous Galaxy. His all-time best view of that galaxy came from an island off the coast of Brazil, where he traveled last summer to begin research on threatened marmosets in the Amazon rainforest. He can be reached at steelshard@tamu.edu. Copyright 1999, 2001 by Astronomical Society of the Pacific, all rights reserved. |